Common trilobite characteristics to help with identification in outcrop and core
Next to the dinosaurs, trilobites are amongst the most recognizable of any group of fossils. They frequently are destined for collections, even with those who have only a passing interest in paleontology. Trilobites define the second stage of the Early Cambrian, beginning 529 million years ago, as full participants in the great explosion of invertebrate critters (the base of the Cambrian is 541 Ma.). They also contribute to the mystery of the Cambrian explosion because they appeared as fully formed arthropods with well-developed segmentation, appendages, complex eyes, and hard protective carapaces. Their precursors probably evolved during the Ediacaran (latest Precambrian) and earliest Cambrian, but what form these ancestral animals took is a problem that awaits solid resolution.
Trilobites are one of the most diverse of all fossilized invertebrates. The number of species burgeoned through the Cambrian and Ordovician, but by the Devonian were depleted by successive extinction events, niche competition, and predation by voracious Placoderms (common Devonian bony fish). The great Permian extinction saw their last gasp. Trilobites were one of the few invertebrate groups to survive almost the entire Paleozoic.
Trilobites secreted a calcium carbonate carapace – this is the preservable part that provides us with a faithful copy of their anatomy. It seems that, like many other arthropods, they shed this carapace at regular intervals (also called molting). Thus, a single critter could produce several exoskeletons, increasing its preservation potential. Overall lengths range from a few millimetres to about half a metre, but most were 5-8 cm long or less.
Most trilobites crawled along the sea floor, probably scavenging for food along the way. With so many legs, all working in unison, it’s not surprising that they left evidence of their activity as fossil traces. A common trace is Cruziana, that consists of bilobed, herringbone-like grooves excavated by the movement of pairs of legs.

Other sources of information
American Museum of Natural History, Trilobite Website
Sam Gon’s excellent website – A Guide to the Orders of Trilobites provides lots of detail
Image credits: Several of the photographs were taken and generously donated by Annette Lokier, University of Derby (indicated on each image).
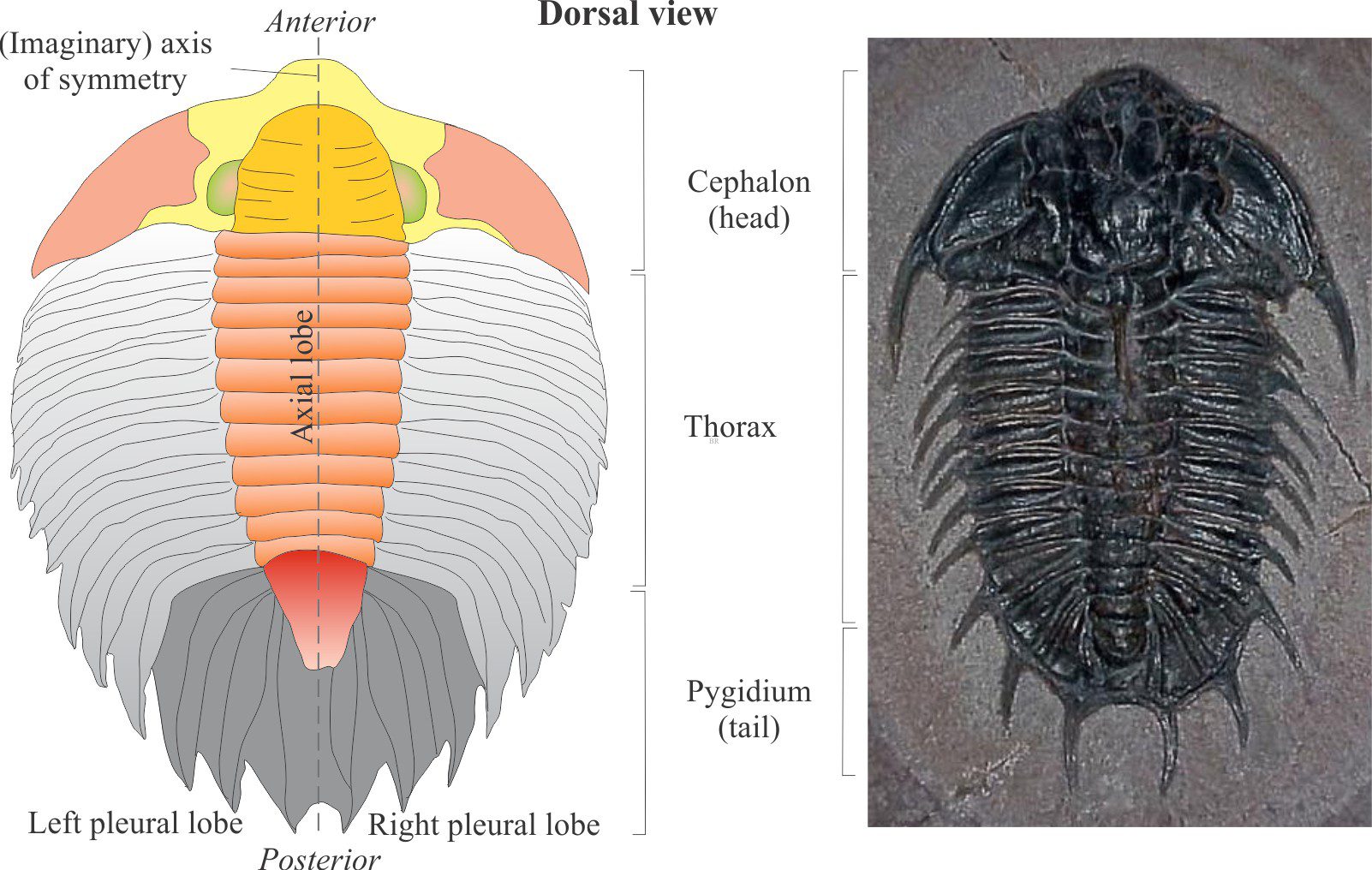
Trilobite orientation
The upper surface that contains the carapace is dorsal; this is the view we usually see in outcrop and hand specimen. The softer underside is ventral; in life this was the side closest to the substrate. The tail (pygidium) margin is posterior and the head (cephalon) margin anterior.
The tri-lobed character of trilobites is defined by a central, longitudinal axial lobe flanked by two marginal or pleural lobes. Thus, there is an (imaginary) axis of bilateral symmetry through the centre such that one half of a trilobite is the mirror image of the other. These are designated left and right pleural lobes when viewing a specimen with the tail down (or closest to you) and anterior up.
Carapace morphology
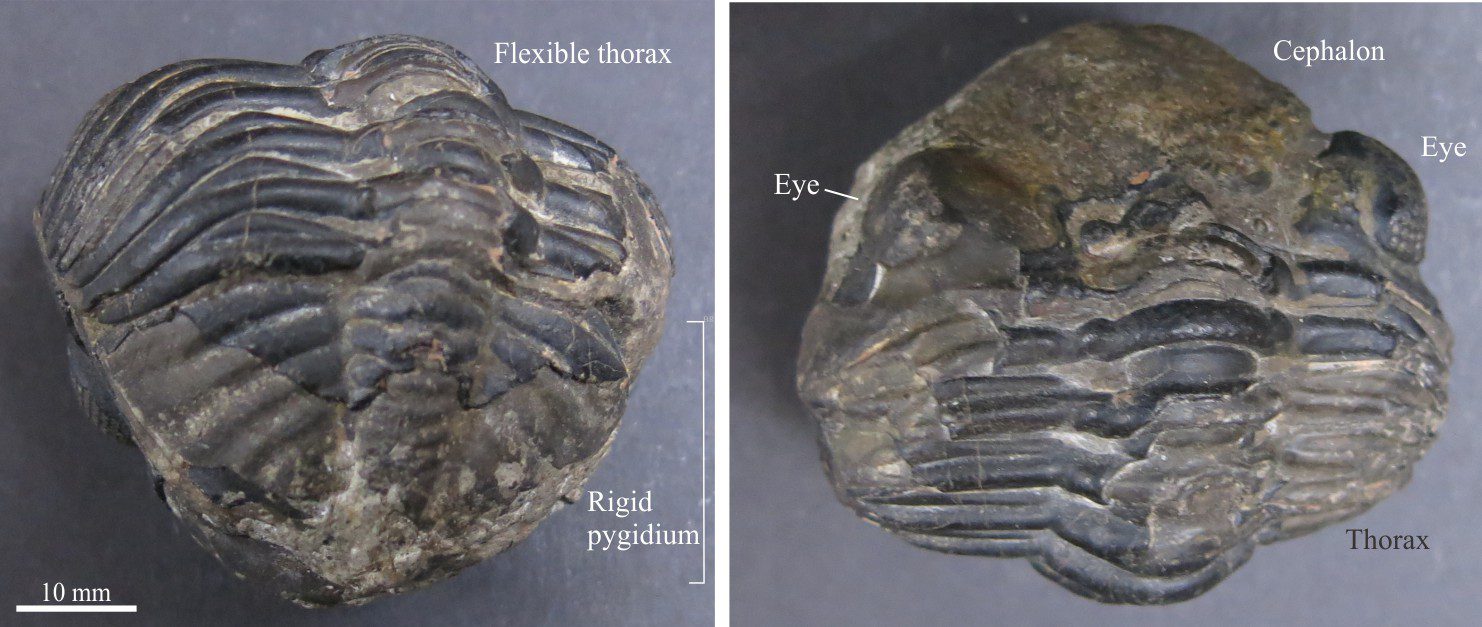
All arthropods are segmented. For the trilobites this includes a cephalon (the head), thorax, and pygidium (equivalent to an abdomen). Each is segmented, although in the cephalon and pygidium these segments are fused together. Segments in the thorax were moveable providing flexibility as the animal crawled over uneven substrates and obstacles or rolled into a ball for protection.
Trilobite heads, from one taxon to another, are much the same size (about a third to a quarter the total trilobite length) and are generally wider than the thorax. Pygidia on the other hand vary considerably in size relative to the cephalon, ranging from millimeters to larger than the associated cephalon. Some examples are shown below. At one extreme, the genus Agnostus has subequal pygidium and cephalon (referred to as isopygous) – it also has only 2 segments in its thorax – apparently Agnostus was blind). In contrast, the common Ordovician genus Calymene had a much smaller pygidium (it is micropygous).

Cephalon
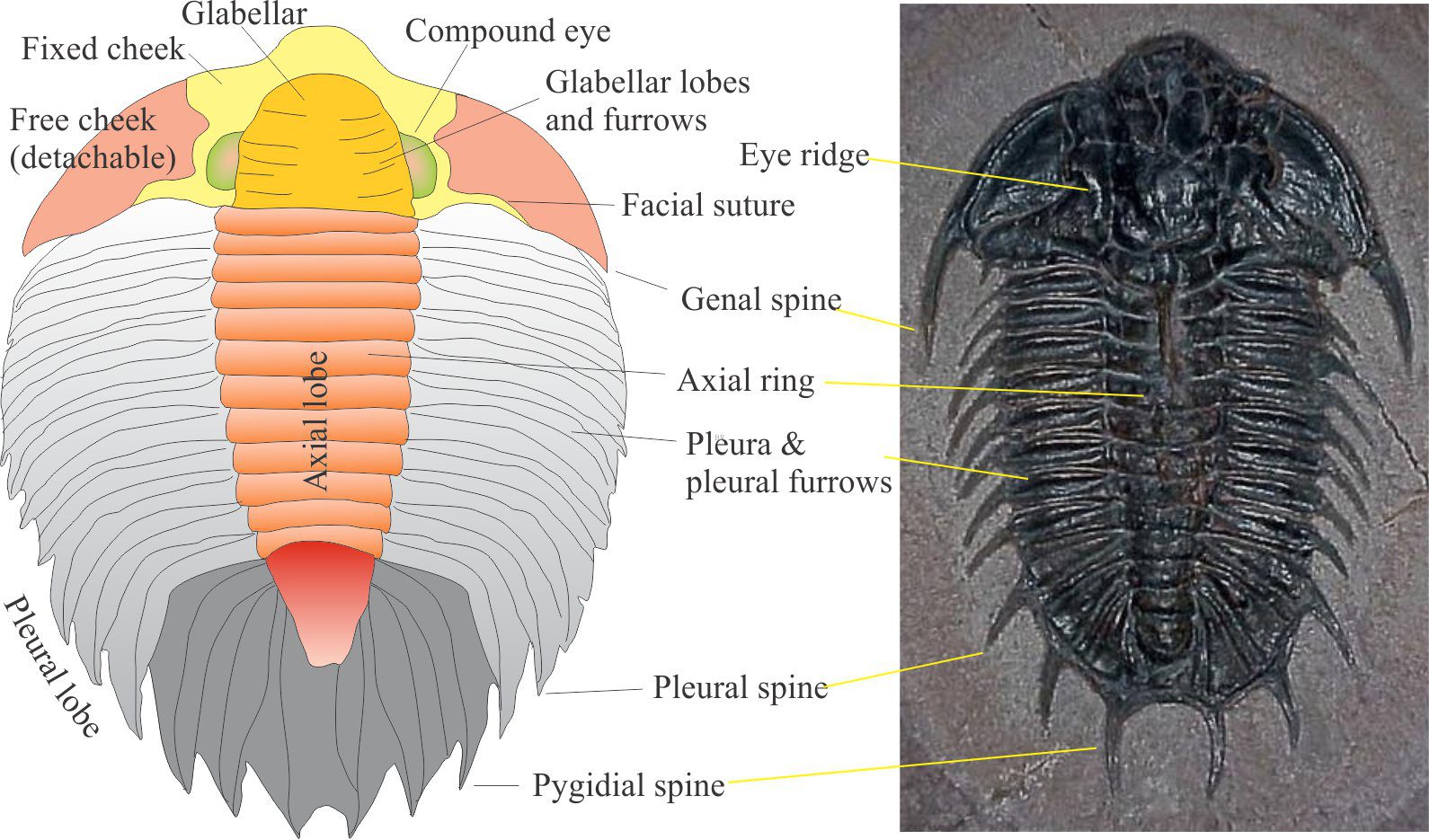
Commonly recognized features of the cephalon include:
- The facial suture which allowed the main part of the carapace to separate from the head during molting. That part of the carapace outside the facial suture – the free cheeks – could be detached, leaving the fixed cheeks
- A central lobe oriented longitudinally called the glabella, that divided the head bilaterally.
- Compound eyes on either side of the glabella. Trilobite eyes were some of the most sophisticated ever to evolve in arthropods. Each eye contained multiple lenses (hundreds of lenses in some taxa), each lens consisting of a single, prismatic calcite crystal. The geometrical disposition of the lenses permitted an almost 360° field of view.
- Genal spine: Usually at the posterior, outer edge of the cephalon, as part of the free cheeks. Spines may be short and stubby or extend almost the entire length of the thorax.


Thorax
This is the flexible part of the animal made up of overlapping segments, or pleurae, each attached to an axial ring in the central axial lobe. Linkages between the cephalon and pygidium were also flexible.
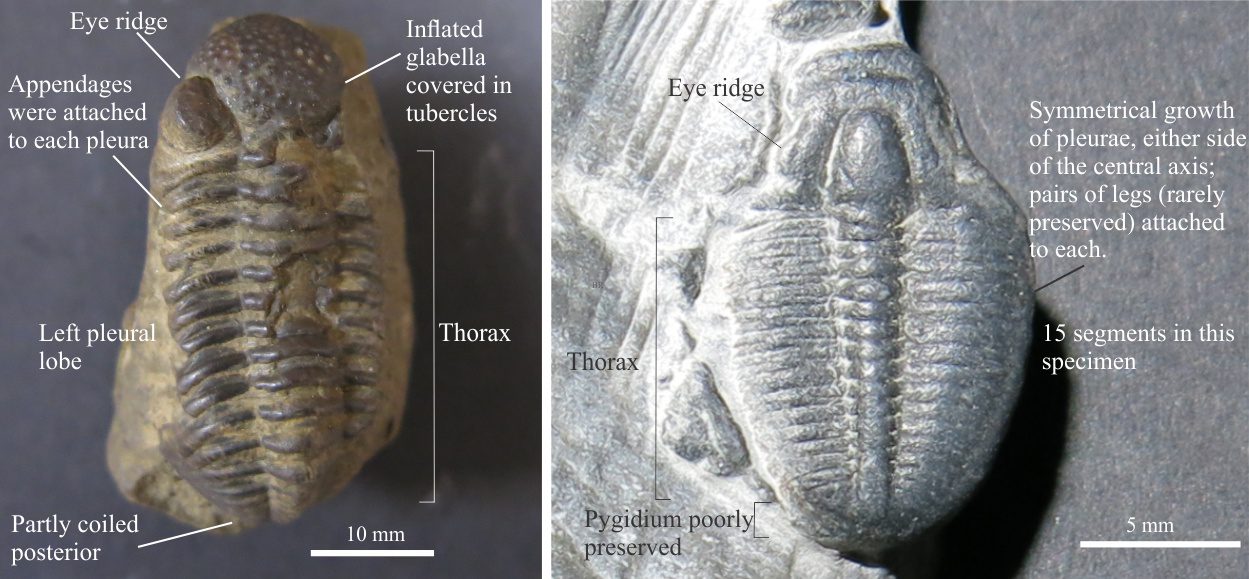
A complete thoracic segment consists of pleura on either side of the axial lobe. Each segment contained a pair of legs, protected in part by the plurae; trilobite legs were never calcified and hence rarely preserved. The genus Agnostus has only two segments. Many species have 10-15 segments like the examples illustrated here; a few have several dozen segments.
The outer margins of plurae frequently culminated in spines. There is still debate over their purpose – were they grown for protection, or to assist with locomotion? In some species the spines also grew from the upper surface of the thorax, rather than the margin.
Pygidium
The tail end of the carapace also consists of pleural and axial ring segments but they are fused1 and therefore inflexible. These pygidial segments also contained pairs of legs that extended from beneath the carapace. The outer margins of each segment were frequently adorned with spines.
Other posts in this series
Bivalve shell morphology for sedimentologists
Gastropod shell morphology for sedimentologists
Cephalopod morphology for sedimentologists
Brachiopod morphology for sedimentologists
Echinoderm morphology for sedimentologists
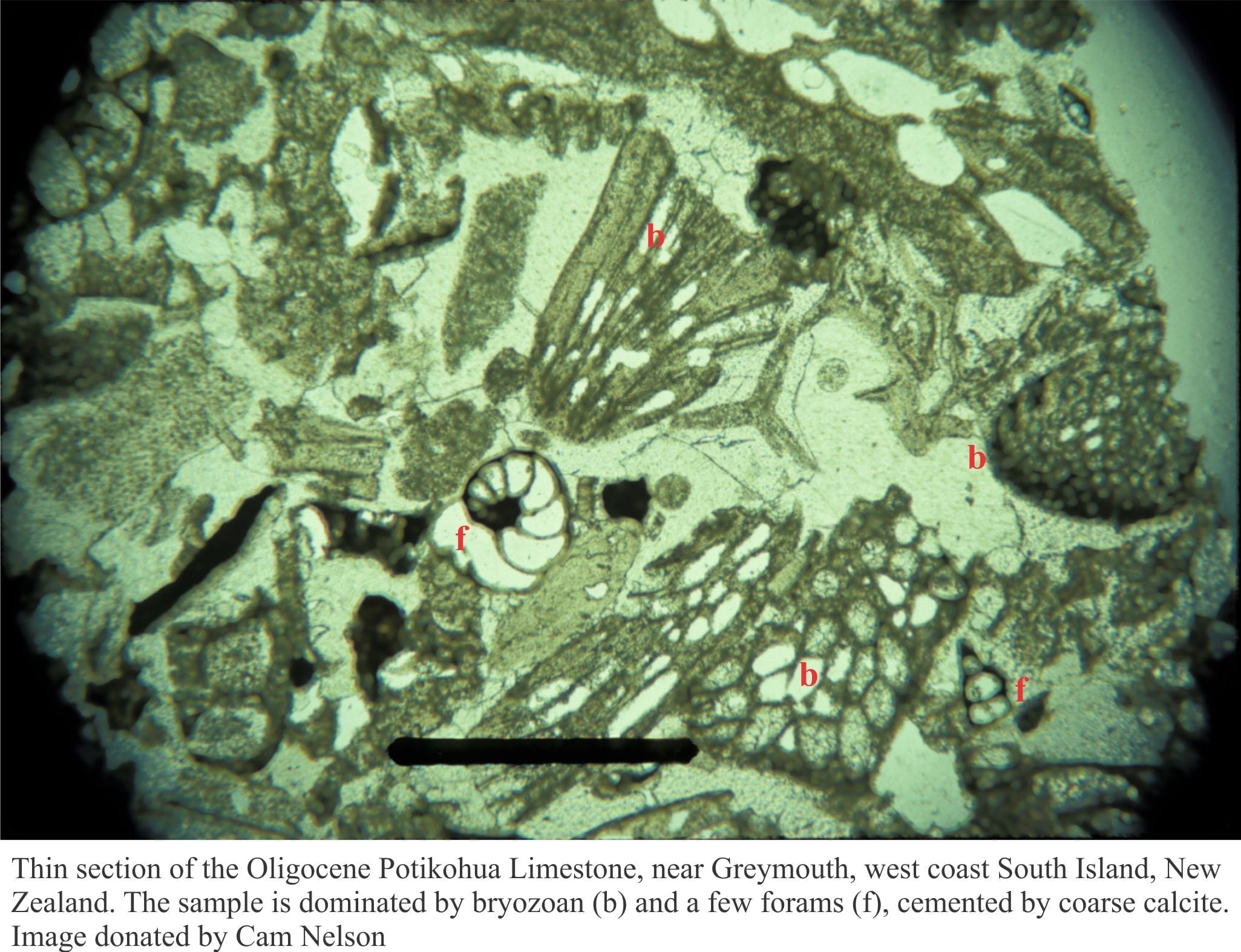
Carbonates in thin section: Forams and Sponges
Carbonates in thin section: Bryozoans
Carbonates in thin section: Echinoderms and barnacles
Carbonates in thin section: Molluscan bioclasts
Neomorphic textures in thin section
Mineralogy of carbonates; skeletal grains