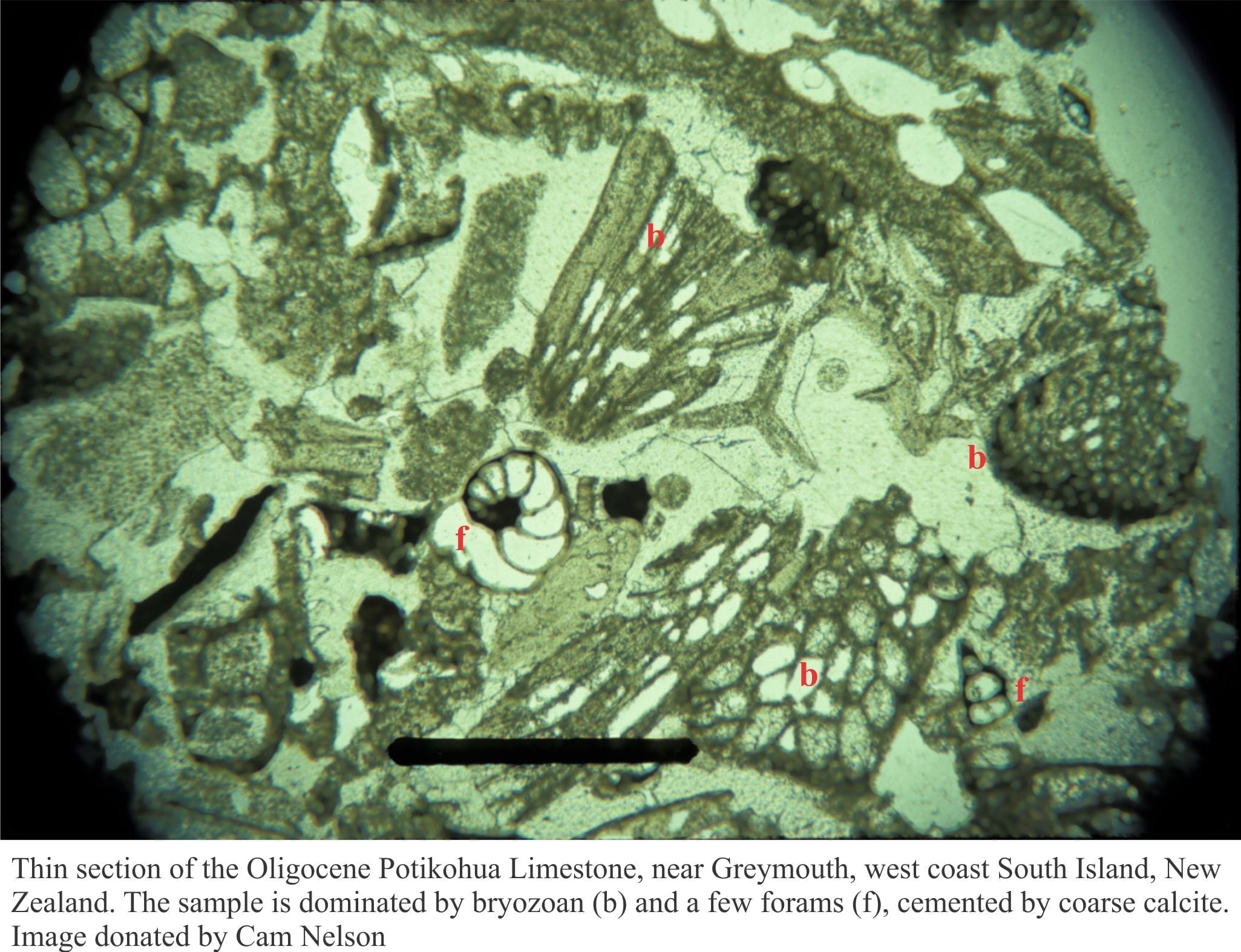
Some criteria that help us identify fossil seagrasses in the rock record
[This post is a companion to Seagrass meadows and ecosystems]
Coastal seagrasses along with mangrove communities are two of the most productive marine ecosystems in the world. Their primary ecosystem functions have been summarized in a companion post. Despite their importance in modern ecosystems and their presumed contributions to fossil depositional systems, they are poorly represented in the actual rock record. There’s a perfectly logical reason for this – seagrasses are angiosperms, flowering monocotyledons that produce leaves, roots, and rhizomes, all of which are readily biodegradable. Seagrasses have very low preservation potential.
So how is it that seagrass communities are presumed to have been important in Late Cretaceous to Recent coastal paleoenvironments? In a few reported cases there is reasonable direct evidence in the form of leaf-rhizome moulds, casts, and bioimmured fragments. But the main lines of evidence are derived from biofacies associations – the array of epifauna and infauna, motile and sessile invertebrates, protozoa (especially foraminifera), and marine algae (particularly those that secrete calcium carbonate) that indirectly indicate seagrass involvement.
Modern seagrass communities thrive in intertidal and shallow subtidal waters, occurring along most of the world’s coasts from the Arctic-Antarctic circles to the tropics. Species diversity is highest in the tropics. Temperate and cool water coasts have fewer species but those that do exist can develop meadows covering many square kilometres (6 species on the Atlantic and Pacific coasts of Canada; 7 in the Mediterranean, 27 in Australia, and a single species of Zostera along New Zealand coasts). This bioregionalism also means fundamental differences in associated lithofacies and biofacies between tropical and cool-temperate environments.

In cool and temperate seas, seagrasses are associated predominantly with heterozoan organisms (those that feed on organic matter), phototrophic algae (such as calcareous red algae) and a variety of invertebrates. These associations exist in the modern tropics (with the addition of calcareous green algae), but they share their habitat with or are marginal to coral reefs. Seagrass communities are also regarded as important components of ramp and platform carbonate factories – primarily because of epiphytic organisms such as encrusting foraminifera, bryozoa, and calcareous red algae, and their function as sediment binders and trappers (e.g., Mazarrasa et al., 2015, PDF).
Recognizing ancient seagrass communities
Most publications that identify paleo-seagrass communities do so using indirect criteria; a selection of these papers is linked herein. In a very few cases there is direct evidence where seagrass leaves and rhizomes are preserved intact or as casts, moulds, or impressions. Reich et al., (2015) provide an excellent review of the criteria for seagrass lithofacies recognition, where the evidence is classified as direct (and reliable), or indirect and less reliable (they use terms like “suggestive” and “weak”). Most of the criteria listed by Reich et al., fall into the indirect categories (see their Table 2) – the criteria tend to be weaker because most of the associated invertebrate, protozoa, and algae biofacies are not unique to seagrass communities.
There are some excellent examples of seagrass leaf and rhizome preservation from the Pliocene of Greece and Canary Islands – examples like this are particularly useful for ‘calibrating’ associated fossil biofacies and lithofacies components for application to the more common circumstance where plant fossils are absent.
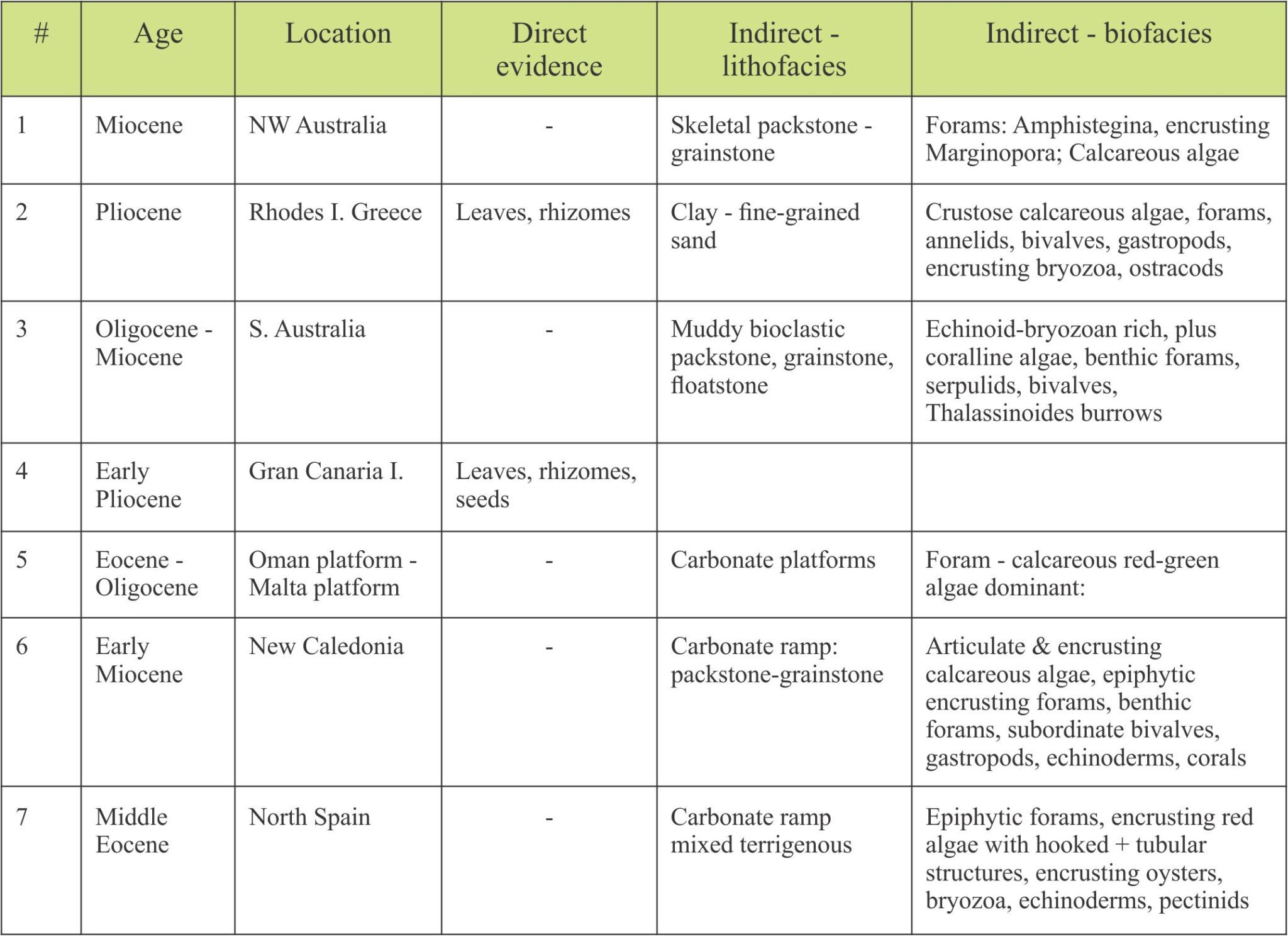
The examples listed in the table below (certainly not exhaustive) illustrate the range of lithologies and biofacies associations that are used to infer fossil seagrass assemblages.
Direct evidence
Although rare, seagrass leaves and roots are documented as carbonaceous remains and as impressions in silicified sediment in rocks as old as Campanian. They are also recognizable as bioimmured casts and micritic linings. The degree of confidence with these interpretations is boosted if leaves and stem are attached, and where epifaunal organisms such as bryozoa and calcareous algae are preserved on the leaves. However, it is possible to confuse these fossilized forms with other coastal terrestrial plants or marine macroalgae.
Well preserved fossil leaves and rhizomes have been reported from Pliocene deposits on Canary Islands and Rhodes Island, Greece. The Gran Canaria site also contains seagrass seeds of the species Halodule (Tuya et al., 2017). The Rhodes Island site contains fossil Posidonia oceanica and a rich assemblage of skeletal hydrozoans and invertebrates including crustose coralline algae, foraminifera, annelids, gastropods, bivalves, encrusting bryozoans, and ostracods (Moisette et al., 2007).
Bioimmuration
Substratum bioimmuration is the process where the skeletal or encrusting material (commonly calcium carbonate) overgrows another organism. This is particularly important where the substrate is otherwise poorly preserved; for example, plant material or soft-bodied invertebrates. There are a variety of organisms that perform this task – encrusting bivalves such as oysters, bryozoa, crustose calcareous algae, the basal plates of barnacles, and corals. The process has the potential to preserve fine details of the substrate structure – in the case of seagrass, this includes leaf branch nodes, leaf veins, and structures on emergent stems.
Indirect associations
Published interpretations of fossil seagrass communities commonly rely on indirect evidence – the presence (or absence) of associated epifauna on leaves and stems, and infauna that roam, graze, or scavenge the sediment-water interface or dwell within the substrate. Epifaunal and infaunal foraminifera figure prominently in these analyses, commonly in tandem with calcareous algae and bryozoans. Most other invertebrates (e.g., bivalves, gastropods, echinoderms, brachiopods, barnacles, crustaceans) may be important contributors to seagrass ecosystems, but they are not diagnostic as such.
Two structures that improve the confidence with which fossil seagrasses can be identified are:
- Hook-like structures formed where the encrusting carbonate extends over a leaf edge – they are best observed in thin sections and polished rock slabs, and
- Tubular structures where a leaf or stem is completely encased by the encrusting organism. In this case care must be taken to distinguish the encrusting form from superficially similar structures like Serpulid (worm) tubes.
Foraminifera
Motile (can move under their own steam) and sessile forams are common cohabitants with seagrass and in the rock record are the most common group of organisms taken to indicate fossil seagrass communities. This applies particularly to species that are permanently attached to leaves and stems; common examples include the genera Sorites, Planorbulina, and Gypsina . Encrusting species may be preserved attached or bioimurred to seagrass leaf casts. In most cases, preservation potential is high, but a recurring problem is that most attached species also occur on other substrates (e.g., macroalgae, coralline algae), and motile species like Amphistegina and Elphidium are common in other intertidal and subtidal environments.
Calcareous algae
Red algae commonly attach to or encrust seagrass leaves and exposed rhizomes, as they do on other substrates not associated with seagrasses such as brown macroalgae (common seaweeds), mollusc shells, coral rubble, and rock fragments (e.g., Lithothamnion). Crustose species are more likely to preserve seagrass leaves via bioimurration. Hook-like structures formed at leaf crust-overhangs are commonly identified as seagrass leaf artifacts.
Articulate coralline algae (branched, flexible) are more commonly associated with seagrasses than other non-vegetated environments, but even this association is not exclusive. Articulate species are also more likely to detach from the leaf substrate during deposition. Articulate and crustose calcareous red algae are important in both tropical and temperate-cool environments, but may be subordinate to calcareous green algae in tropical settings (e.g., Halimeda, Penicillus).
Calcareous green algae such as Halimeda and Penicillus are photosynthetic, tropical marine dwellers. They are important contributors to carbonate mud factories. They associate with seagrass communities on carbonate platforms and the shallower margins of carbonate ramps, and with a variety of coral reefs in adjacent lagoons or shallow forereef sites.

Bryozoans

Bryozoans are a diverse phylum in many shallow marine environments, a diversity that also applies to seagrass communities where both calcareous and non-calcareous species thrive (>150 bryozoa species are associated with some Mediterranean seagrasses; Reich et al, op cit.). Bryozoans that encrust seagrass leaves tend to be unilaminar and less permanent than species that grow on stems and exposed rhizomes – because of leaf flexibility. Colonies that grow on leaves also tend to expand along the length of the leaf and can completely envelop leaves and stems. There may also be evidence for bioimurration of seagrass leaves and rhizomes by encrusting bryozoa (e.g. Taylor and Di Martino, 2014, PDF).
Other invertebrates (bivalves, gastropods, echinoderms)
Infaunal bivalves in many tropical and temperate seagrass communities include the common suspension and deposit feeders (e.g. Venerids, Carditids, Tellinids, and Nuculids), but most of these species also thrive in non-vegetated shallow marine environments. Likewise, epifaunal communities commonly include oysters and Pectinid species, but these too are wide-ranging in other habitats. The mussel family Pinnidae (fan mussels) are common in seagrass substrates but also extend to non-vegetated environments. A similar situation exists for gastropods, although the species Smaragdia may prefer seagrasses. Echinoderms are common inhabitants of seagrass meadows – some species graze on the leaves – but most extend to other non-vegetated settings.

Trace fossils
Given the biological vitality of seagrass communities in intertidal and subtidal environments, it is not surprising that traces and burrows are common in seagrass substrates – we see this in modern depositional settings and their fossil analogues. Burrowing organisms are numerous in these communities – gastropods and echinoderms, crustaceans (crabs, shrimp, sand hoppers); the traces will reflect their diverse behaviours. Common ichnofauna include Thalassinoides, Ophiomorpha, Skolithos, and Scolicia, However, most of these trace fossils are also common in non-vegetated shallow marine settings, so their potential value as seagrass indicators is limited.