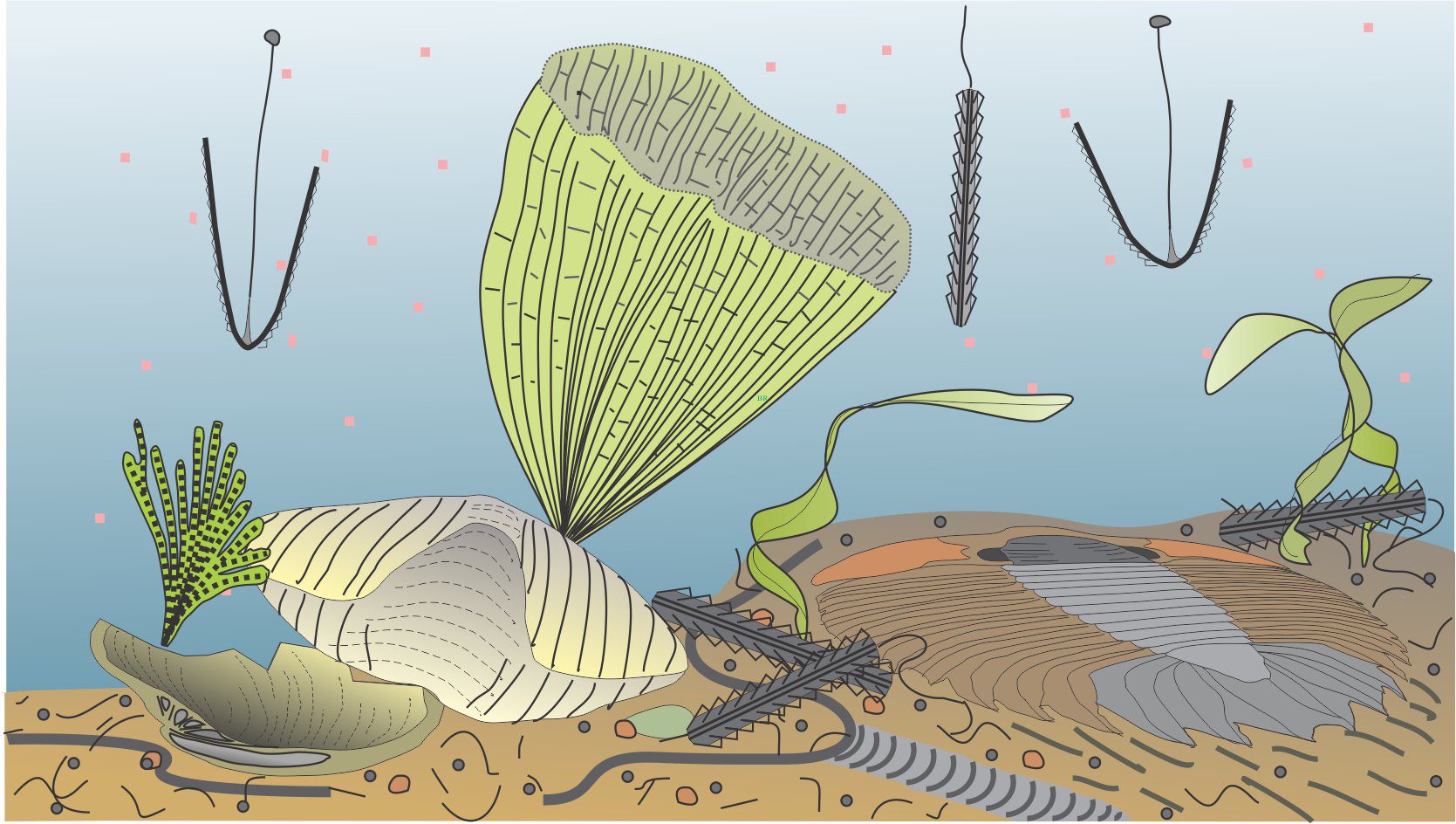
For good reason, Early Paleozoic hemichordate fossils derive their name from the Greek graptos, and the Latin version graptolithus meaning ‘written on stone’; at a superficial level of observation, they look like the scribblings of some errant rock-god. Graptolites are mostly preserved in shale and recovered from rocks that split easily along bedding. And in this state they do appear as doodles or smears – the remains of thriving, colonial communities of marine animals.
Graptolites arrived on the scene in the Middle Cambrian and disappeared toward the end of the Carboniferous. Their preserved morphology and evolutionary links to some modern hemichordates indicate that they were probably colonial organisms. They are included in the hemichordates because some specimens present evidence of a notochord, a rod-like cartilage supporting structure that maybe the precursor to the cartilage part of a backbone. As such, graptolites are considered possible ancestral candidates to full blown chordates, the phylum that includes vertebrates.
The biostratigraphic value of graptolites was first recognised in the mid-1800s in Canada, Sweden, Bohemia, and Scotland. Charles Lapworth who first identified the inverted stratigraphy associated with what is now known as the Moine Thrust, was one of the first to use graptolites to resolve stratigraphic problems – he also recognised the Ordovician as a geological period separating the Cambrian and Silurian. The biozones identified by Lapworth are still used today, with a few modifications and updates. Their biostratigraphic value lies in the widespread distribution and rapid evolutionary differentiation that allows high resolution stratigraphic subdivision of Lower Paleozoic successions. As an example, see the paper by Cooper and Lindholm (2009) for details of high resolution subdivision of the Ordovician. For a summary of British graptolite biozones see Zalasiewicz et al., 2009 (PDF available).
The specimen photographs shown here were generously provided by Annette Lokier, University of Derby.
Modern relatives
The closest living graptolite relatives are a group of hemichordates – the pterobranchs which are sessile, colonial, worm-like animals constructed by filter-feeding zooids that secrete collagen (a complex protein) or chitin-like tubes. Although the zooids are individual animals, they are interdependent. Recognition of the pterobranch – graptolite relationship is important because graptolites had no hard skeleton and their preservation in 3-dimensions is rare. Graptolite colonies were commonly three-dimensional structures, but we generally see them preserved as flattened, two dimensional specimens.
Graptolite morphology
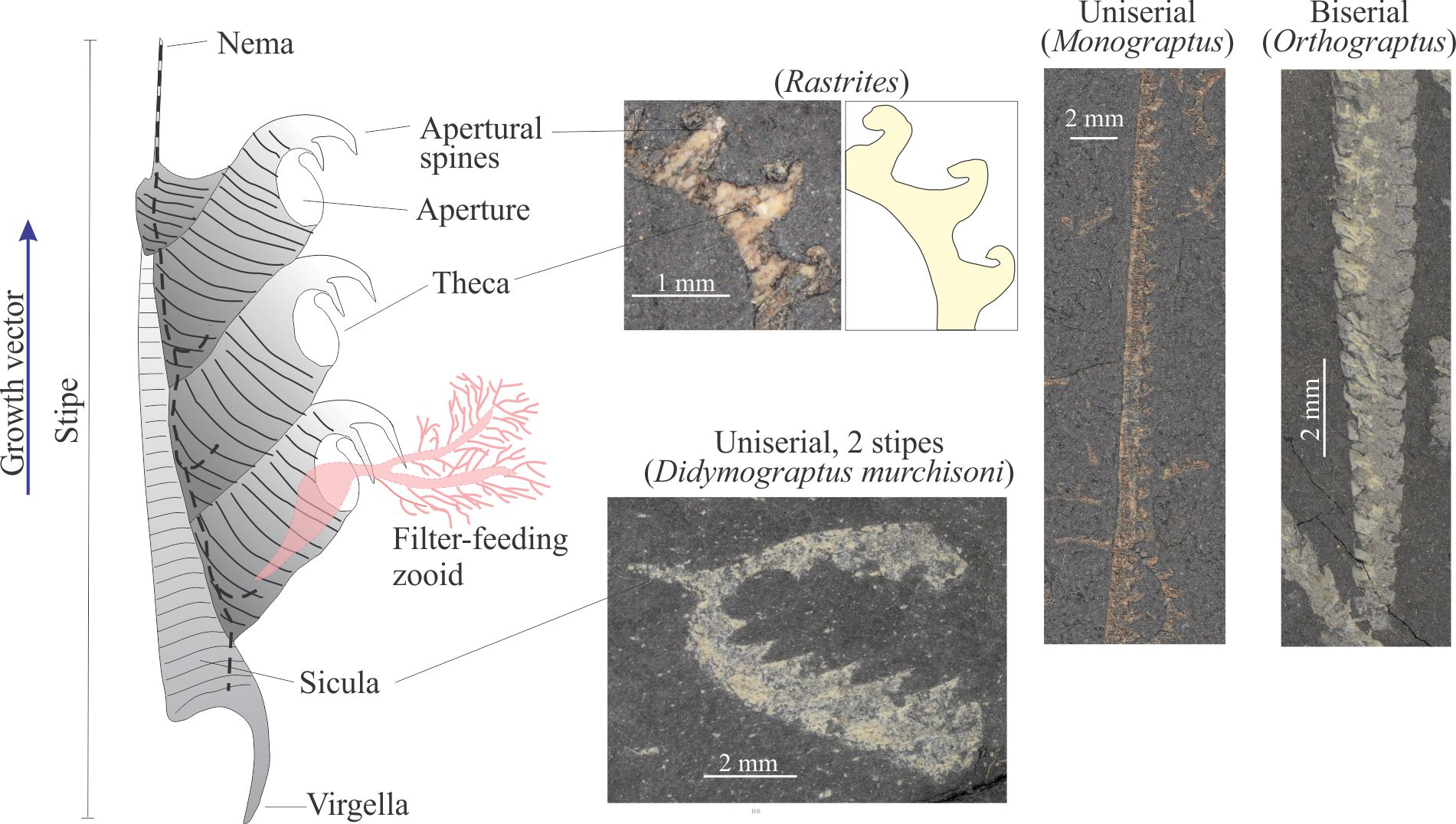
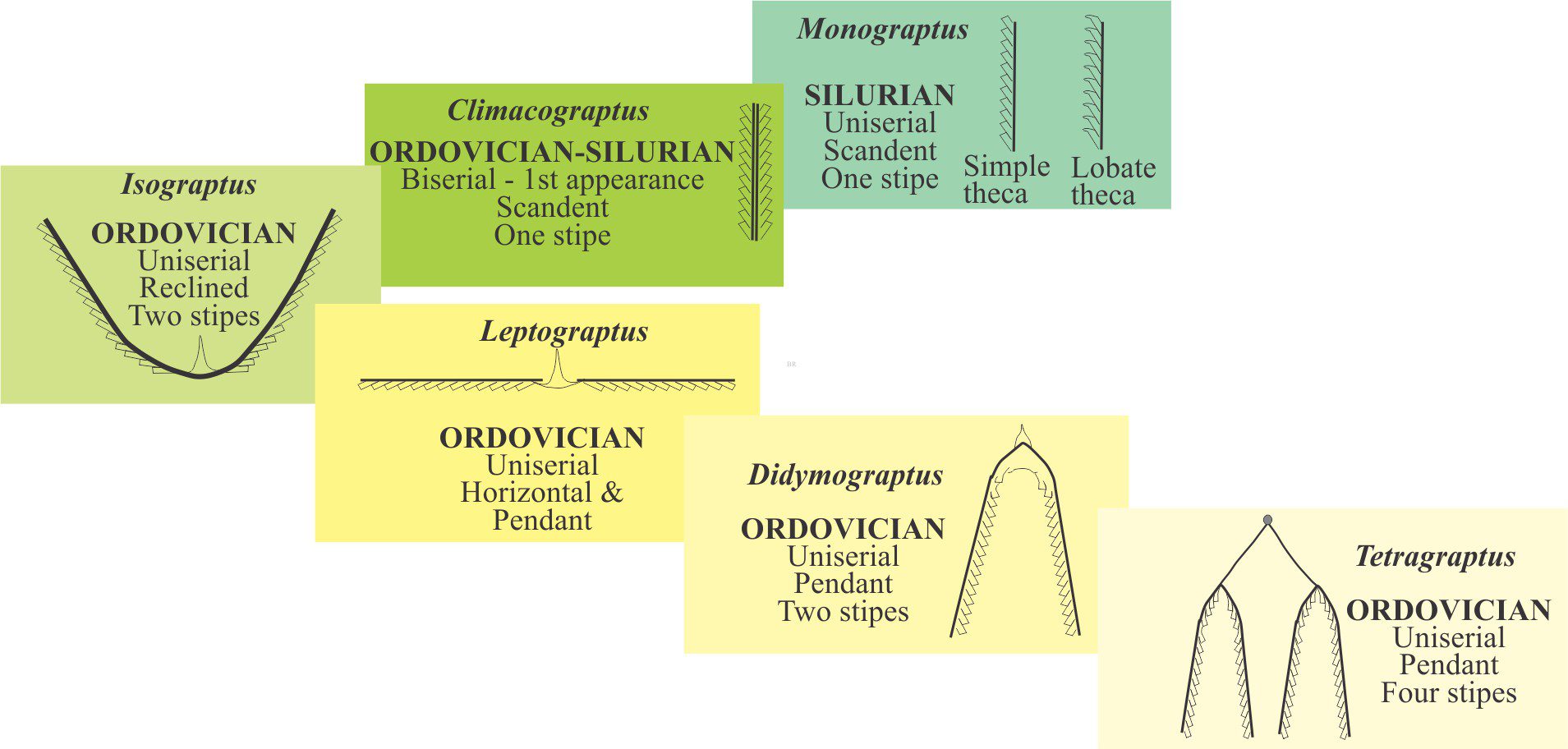
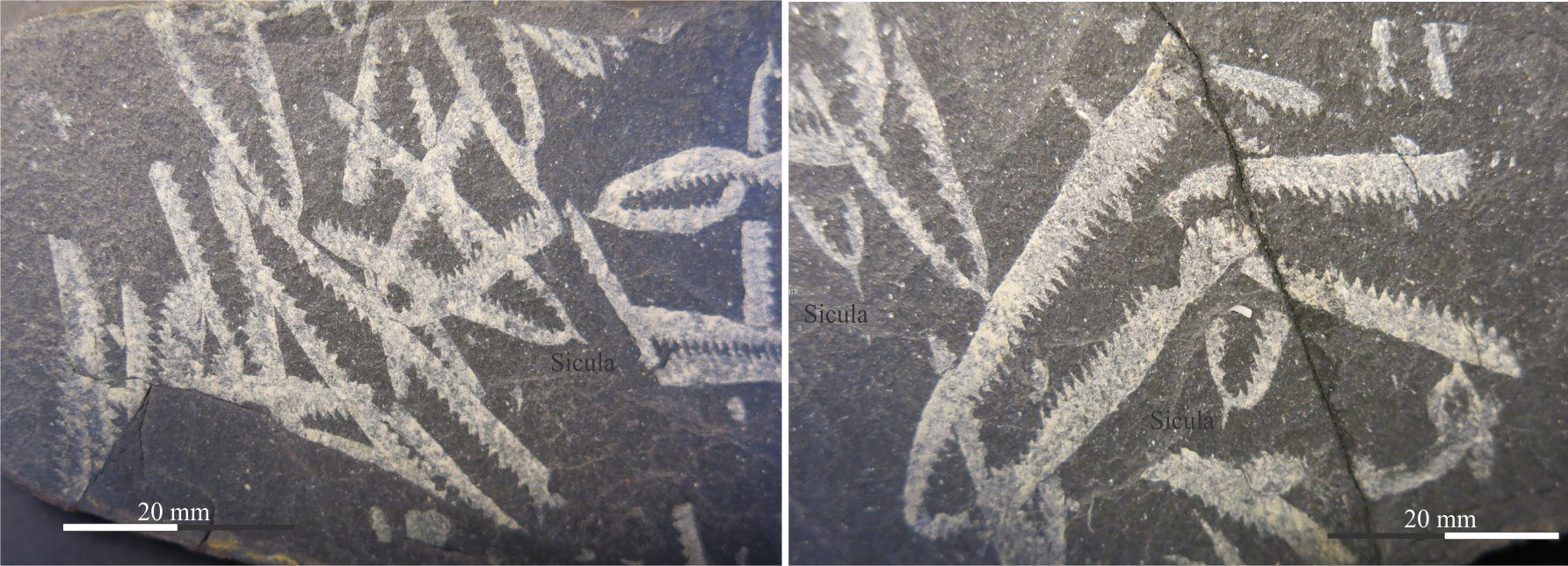
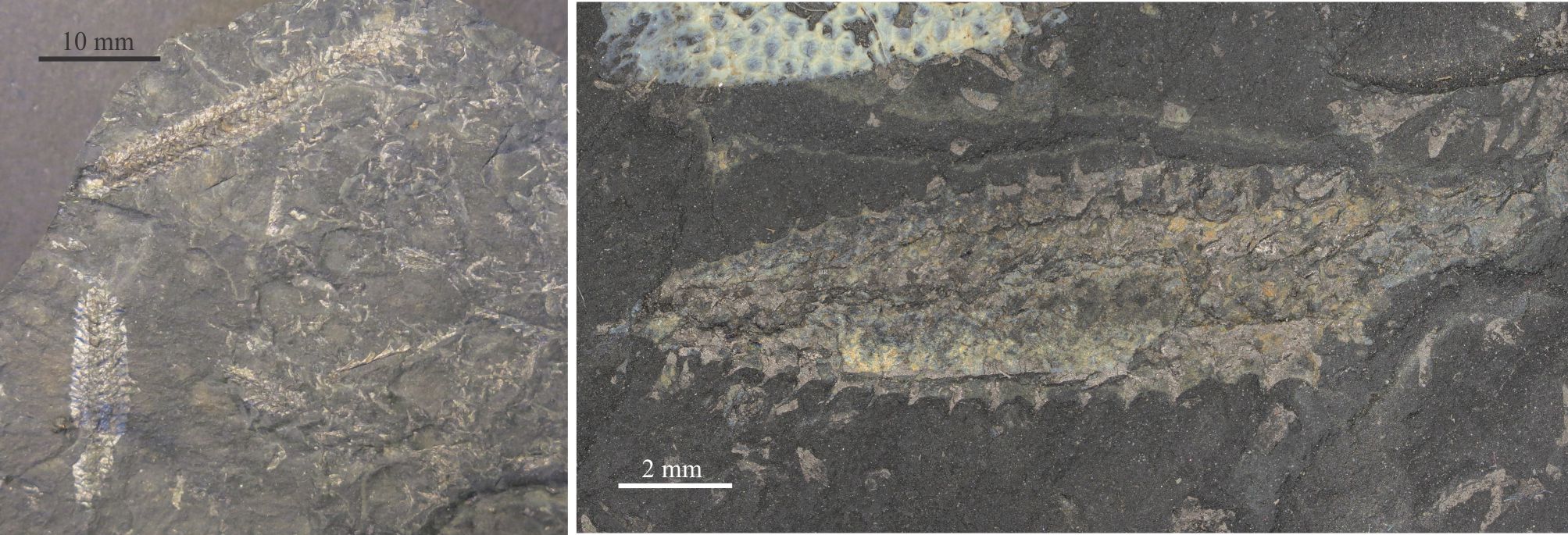
What is known about graptolites has been teased from modern pterobranchs, reconstructions from their 2D fossil preservation in shales, and from those rare cases where 3D form can be deciphered, for example in fine-grained limestone where calcite that encased the preserved animal is dissolved. Thus, it is fairly certain that graptolites were colonial and probably constructed by zooids, individual animals that secreted interconnected tube-like domiciles. The first tube is cone-like (a sicula) from which later tubes (thecae; singular theca) grew in a branch-like succession. Each branch is called a stipe. Stipes may be uniserial with thecae on one side, or biserial where thecae line opposite sides of the branch and share the same nema. Quadriserial forms contain rows of four thecae around stipes that appear to have joined in a central tube; Phyllograptus is a good example of this type (shown below). The entire colony is called a rhabdosome. The sicula end of a stipe is commonly referred to as proximal; the opposite end distal (see Maletz et al., 2014 in the Treatise Online Glossary, for detailed definitions and descriptions – PDF available).
Each theca housed a zooid. In modern pterobranchs the opening at the top of the thecal tube (aperture) allows the zooid to extend its feathery feeding filters. Lobes, spines, and hook-like structures may extend from the aperture margin; they are thought to have helped planktic colonies stabilize their motion while suspended in the water column. A hollow thread, or nema, grew through the centre of each stipe, beginning in the sicula, connecting the thecae and the zooids that lived within. The nema continued to grow as new thecae were added to stipes. On uniserial forms the visible part of the thread commonly extended from the top of the latest theca of the stipe. On multiserial forms the nema commonly extended directly from the sicula, for example on the genus Isograptus (illustrated below). One function of the nema was to attach rhabdosomes (entire colonies) to a float. In many graptoloids another spine-like structure, the virgella grew from and was part of the sicula.
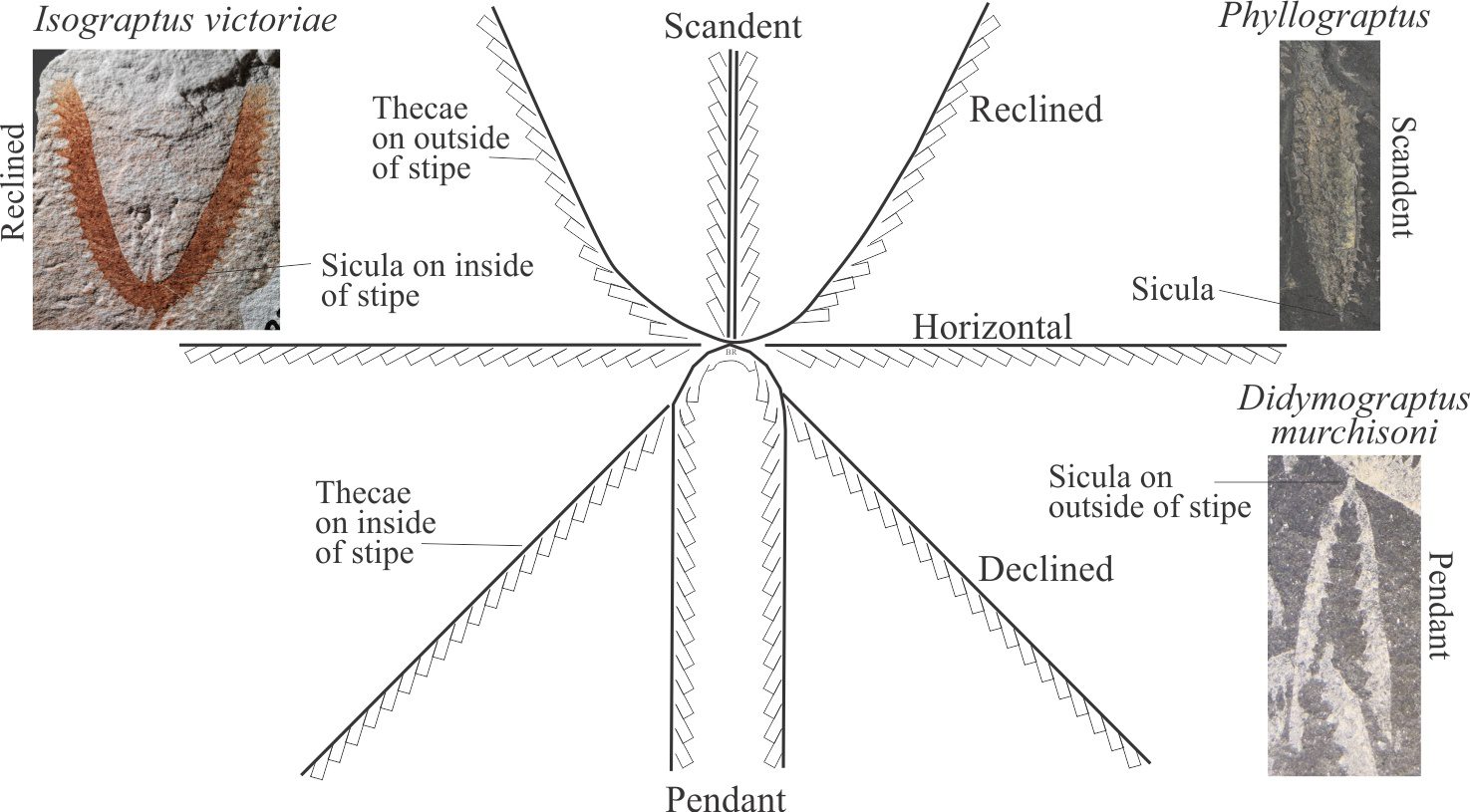
Rhabdosome orientation
Graptolite colonies are defined by the arrangement of stipes and whether thecae are added upwards, downwards, laterally, or obliquely. Note that thecae open towards the nema and away from the sicula. The manner in which successive thecae are added and the overall branching geometry are two of the more important criteria for deciphering evolutionary trends. The most common arrangements are:
- Scandent: Successive thecae are added vertically upward from the sicula, on the outside of the stipes. The examples shown are Phyllograptus, Climacograptus.
- Horizontal: Thecae are added outward (laterally) from the sicula.
- Reclined: Thecae are added obliquely upwards from the sicula on the outside margin of the stipe. The sicula is located on the inside margin of the U- or V-shape stipes. The example shown below is of Isograptus victoriae.
- Declined: Thecae are added obliquely downwards from the sicula (same direction as pendant stipes), on the inside margin of the stipes (i.e., inside the inverted V- or U-shape.
- Pendant: Thecae are added downward from the sicula on the inside margin of the stipe. The sicula is located on the outside margin of the stipes. The example shown is Didymograptus murchisoni.
Some species evolved curved or spiral stipes that provided either greater floating stability or allowed them to rotate in the water column, a strategy that provided greater access to food. Rastrites is a good example.

Evolutionary trends
Recent taxonomic subdivisions define two main orders: dendroids and graptoloids. The earliest representatives were sessile dendroids, appearing in the Middle Cambrian and surviving until the latter part of the Carboniferous (about 320 Ma). Some planktonic dendroids appeared in the Ordovician.
However, the dominant planktonic forms that appeared at the beginning of the Ordovician were the graptoloids. All graptoloids were planktonic although whether they were all free floating or evolved mechanisms to propel themselves through water is still debated. The important differences between the two orders are:
- Sessile dendroids began life with a sicula attached to hard substrates and grew into bush- or fan-like rhabdosomes. Each colony contained many uniserial stipes that were connected by transverse dissepiments. Stipes could also bifurcate. Being sessile, they would have shared the substrates with brachiopods, bryozoa, trilobites, and other benthic organisms. The dendroids were a fairly conservative group with little change in morphology or growth habit for their entire stratigraphic range.


- All graptoloids began life by secreting a floating sicula. Graptoloids consistently grew fewer stipes and mostly lacked the dissepiments and stipe bifurcations that were characteristic of the dendroids. Structures like the nema and virgella are prominent in the graptoloids. Unlike the dendroids, graptoloids evolved a range of morphological forms, ranging from colonies with a single stipe, to those with four stipes – summarised in the diagram below. They also evolved uniserial, biserial, and quadriserial stipes (the dendroids were uniserial). Graptoloids were probably the first major planktic forms to occupy Early Paleozoic oceans, taking advantage of all the nutrients available throughout the ancient water columns. They also became widely distributed. The Ordovician and Silurian periods witnessed the acme of graptoloid evolution – hence their great value as biostratigraphic indicators. The overall evolutionary trend was towards fewer stipes, culminating in single stipe forms like Monograptus. One important evolutionary change in the Middle Ordovician was the appearance of scandent biserial forms like Climacograptus. Graptoloids died out by the Middle Devonian, about 65 million years earlier than the dendroids.




Other posts in this series
Bivalve morphology for sedimentologists
Trilobite morphology for sedimentologists
Gastropod shell morphology for sedimentologists
Cephalopod morphology for sedimentologists
Brachiopod morphology for sedimentologists