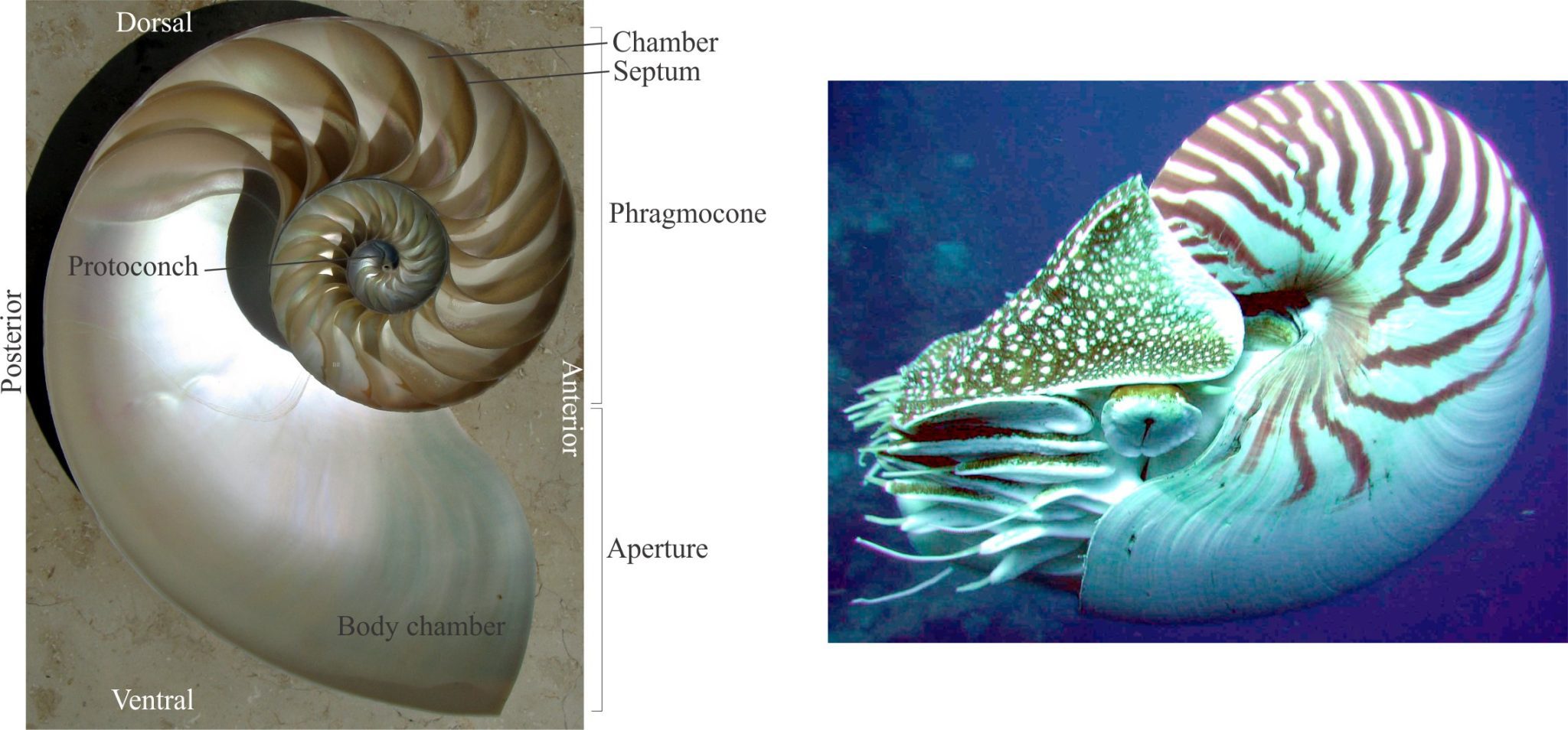
Our knowledge of how ammonoids and nautiloids functioned is based almost entirely on a single extant species – the chambered Nautilus.
What better way to introduce a post on cephalopods than with an image of a very tidy Mary Anning (shoes and skirt hems unmuddied), pointing knowingly at an ammonite. Knowing, perhaps because she had collected the sample, had identified its taxonomy and stratigraphic relevance, and that she might be called upon to explain all this to renowned geologists like Robert Murchison or William Buckland who in turn would largely ignore her contributions to the broader scientific community.
Anning’s ammonites, found in Jurassic strata exposed along the Lyme Regis coast (south England), along with their Cretaceous descendants, represent the acme of cephalopod evolution. They and other cephalopod groups became extinct during the terminal Cretaceous, about the same time as the dinosaurs. Today, surviving cephalopods (about 800 species) are represented primarily by squids, octopuses, and cuttlefish that either lack shelly structures completely or retain them internally. Straight-shelled belemnites are familiar but extinct Mesozoic fossils that represent the internal shells of ancient squid-like critters. The extant coiled ‘ram’s horn’ shells Spirula spirula, that commonly wash in with the tide, are examples of modern shells that provide internal support to a species of squid. The only modern survivors of the external, coiled cephalopods are a few species of Nautilus.
[According to various etymologies, Octopuses is the correct English noun plural for the original Greek plural octopodes, rather than octopi which incorrectly uses the suffix ‘i’ for a word not originally of Latin origin]
Other sources of information
There are lots… but here are a few links to recent texts.
The Cephalopod Page the personal web page of Dr. James B. Wood’s since 1995. A great resource.
The Paleontological Society provides free access to its Digital Atlas of Ancient Life that contains oodles of explanatory texts, field guides, Apps, and images on the fossil record.
Bringing Fossils to Life: An Introduction to Paleobiology, Donald Prothero. Now in its 3rd Edition.
Modern cephalopods
It is thought that many of the characteristics of modern cephalopods also applied to their ancient cousins.
All modern cephalopods are free swimmers and hence are different from all other molluscs. A jet-like structure near the ventral margin of the aperture (below the head) is used for propulsion. The common squid and octopus have 8 arms and a pair of retractable tentacles; suckers are commonly arrayed along each arm. The external-shelled Nautilus has up to 90 arms.
Modern cephalopods have a well-defined head with an oversized brain (compared with other invertebrates), clear evidence of intelligence, particularly in the octopuses, a well-developed eye, and a mouth. They are, and were predatory, eating crustaceans, small fish, and other cephalopods; a radula, analogous to those found in gastropods, assists with chopping up their prey. Modern species have perfected the art of camouflage by changing colour rapidly. Some of the largest squids measure in at 13m long; the smallest octopus is only 2-5 cm. Giant squid-like creatures figure prominently in mythology and literature – witness Captain Nemo’s battle in Jules Verne’s Twenty Thousand Leagues Under The Sea.
Familiar fossil groups like the (almost extinct) nautiloids and fully extinct ammonoids had external shells. Our understanding of their shell structure and life habit derives primarily from observations of the single extant nautiloid genus – Nautilus.
The Nautilus analogy
Year after year beheld the silent toil
That spread his lustrous coil;
Still, as the spiral grew,
He left the past year’s dwelling for the new,
Stole with soft step its shining archway through,
Built up its idle door,
Stretched in his last-found home, and knew the old no more.
3rd stanza from “The Chambered Nautilus“, Oliver Wendell Holmes 1858 poem that celebrates life, death, and the human condition.
The nautiloids were the earliest group of cephalopods, appearing in the Cambrian initially as straight-shelled forms (orthocones), evolving to coiled structures in the Ordovician. Most of the attributes described here apply to nautiloids and ammonoids.
[Fibonacci brilliantly teased the logarithmic sequence while trying to solve a problem involving rabbit breeding – we also see these sequences in sunflowers, hurricane-cyclone cloud patterns and galaxies]
As the animal grows, chambers are added along a plane of symmetry resulting in planispiral coiling, similar to some gastropods. Shell growth is logarithmic, which means that successive chambers are larger than previous chambers. Chambers are separated by solid transverse walls, or septa. Nautiloid septa are relatively simple curved planes. The trace of the intersection of septa with the shell wall defines sutures that in nautiloids trace as simple curves, and in the ammonoids assume more complex geometries.
The animal occupies the latest constructed chamber – the body chamber that opens at an aperture. All formerly occupied chambers constitute the phragmocone. Previously occupied chambers are abandoned by the animal but remain interconnected by a thin, continuous tube – the siphuncle which contains a siphon that allows the animal to regulate the fluid and gas content of each chamber, that in turn controls shell buoyancy and orientation. The siphuncle is held in place by shorter calcareous tubes, or septal necks that are cemented to the septa. In nautiloids the siphuncle is in the centre of each septum; in the ammonoids it occurs along the outer margin of each coil. Shells are composed of two layers of aragonite, the inner layer presenting a characteristic nacreous lustre.
Orientation: In life, the shell is upright with the plane of symmetry vertical and the aperture downward. When the aperture opens toward an observer, the margin facing is anterior and that opposite is posterior. Along the planispiral plane of symmetry, the dorsal margin is upper, and the lower margin is ventral. If the outer margin of a coil or chamber is convex it is called the venter, if concave it is the dorsum.
Two additional terms are used to describe the orientation of shell structures such as sutures and ornamentation, with respect to the aperture, or the apex or protoconch (the first chambers – analogous to gastropods); in particular, these terms are important descriptors of suture geometry. They apply to orthoconic (straight) and coiled shells:
- Adapical – oriented towards the apex.
- Adapertural – oriented towards the aperture.
For example, ribs transverse to the direction of coiling are commonly convex toward the protoconch, or adapical.
Spirula spirula
Referred to as ‘Ram’s Horn’ shells, naked Spirula are frequent visitors along Atlantic and Pacific shores. They are the coiled, internal, aragonite superstructure to a species of squid. Their chambers are connected by a siphuncle that extends along the ventral margin. Like Nautilus, the siphuncle acts to modulate buoyancy. The septa are simple concave walls separating each chamber. A septal neck guides the siphuncle from one septum to the next. The shell is quite fragile and breaks easily.

Coiling geometries in cephalopods
Evolution of coiling in nautiloids and ammonites resulted in a variety of interesting geometries, but the most common was tight, planispiral, homomorphic coiling – most of the examples below are of this type. Irregularly coiled taxa are heteromorphs – a good example is Scaphites where initial chambers are tightly coiled, followed by a straighter segment, ending with a loosely coiled hook-like form.
Examples of coiling and other morphological elements are shown below. Most of the photographs were taken and generously donated by Annette Lokier, University of Derby (indicated on each image).
Orthoconic:
Chambers arranged in a straight cone. Early nautiloids and ammonoids were orthoconic. Later forms developed curvature – these are referred to as cyrtoconic. All belemnites are orthoconic.

Evolute coiling:
Successive coils are in contact but there is no or very little overlap.



Convolute coiling:
New chambers partly overlap older coils.

Involute coiling:
The latest part of the coil, including the body chamber, overlaps all previous coils.

Lituiticone coiling:
Coiling is initially tight and involute with later chambers more loosely coiled and hook-like. The irregular coiling is referred to as heteromorphic.

Ammonoides
The first coiled ammonoids appeared in the Devonian. Because they are extinct, any inference about animal structure, life-style, and other paleoecological attributes is largely dependent on studies of modern cephalopods, together with the various sedimentological and paleontological facies with which they are associated.
Ammonoid evolution accelerated during the Mesozoic, culminating in the Cretaceous with a diverse array of coiling geometries and suture patterns. The more common coiling patterns are described above.
Two important shell structures that distinguish ammonoids from nautiloids are:
- The position of the siphuncle, that in ammonoids tracks around the outer margins of chambers (in nautiloids it traverses the centre of successive septa).
- The complexity of their sutures.

Sutures
The intersection of septa with the shell wall is manifested as sutures that are commonly visible on the outer shell surface. In nautiloids and early ammonoids the sutures are usually straight or simple curves. The curves represent the convexity of corresponding septa that usually are concave towards the aperture. However, during the Mesozoic and culminating in the Cretaceous, ammonite sutures became increasingly complex with the development of intricate alternations of lobes and saddles: lobes are adapical (closing towards the apex or protoconch), saddles are adapertural. Lobes tend to be more pointed and have larger amplitudes than saddles (Klug and Hoffmann, 2015 – PDF available – a good review of sutural styles and structures). Lobes and saddles may be complicated by the addition of smaller, secondary folds. Sutural complexity reflects the same degree of complexity in the corresponding septa. A couple of examples are shown below.
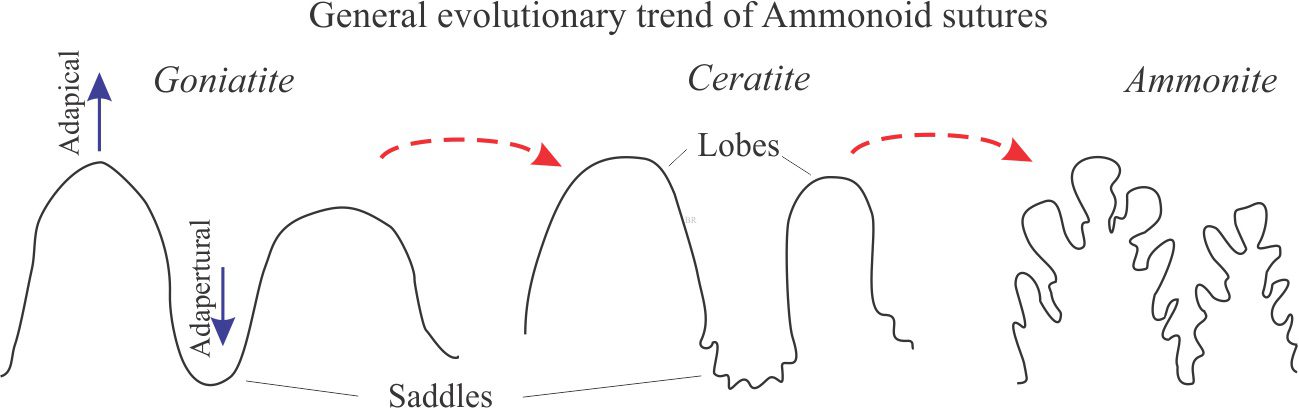
The complexity of ammonoid sutures increased through the Paleozoic, culminating in the remarkably intricate suture patterns of later Mesozoic ammonites. Three suture pattern categories are recognised that are the basis for taxonomic classification into three subclasses: Goniatite sutures, consisting of simple curved sutures and no subdivision of lobes and saddles (see the image of Cheiloceras above) (mostly Devonian to Permian); Ceratite sutures that have lobes subdivided into smaller-scale fold structures (mostly Triassic); and Ammonite sutures characterized by intricately subdivided lobes and saddles (mostly Jurassic-Cretaceous). Two examples of the latter are shown below.


Belemnites
Along with the ammonites, belemnites had their hay day in the latter part of the Mesozoic. They are a common, extinct group of squid-like creatures that taxonomically are slotted into the Coleoidea, a subclass that includes modern squids and cuttlefish.
The shelly parts of belemnites were internal. Their phragmocone was straight (orthoconic), consisting of a simple, conical array of septa, beginning with a protoconch at the pointy end (posterior), and ending with the latest, anterior chamber to which the animal was attached. Septa walls were probably aragonitic. Older chambers were connected by a soft, ventral siphuncle that vented fluid and gas to maintain buoyancy, in much the same way as Spirula spirula and orthoconic or coiled ammonites. Orientation is determined by facing the phragmocone chamber, dorsal is up, and ventral down (same as ammonoids and nautiloids). The phragmocone (that contains the chambers) fits neatly into a solid, bullet-shaped guard, or rostrum that was secreted as radially crystalline, low-magnesium calcite (Hoffmann and Stevens, 2020, Open access).
Although the soft parts of Coleoids have low preservation potential, there are a few examples of exquisitely preserved belemnites that show an array of ten arms, each armed with as many as 40 hook-like structures that probably were used to catch and hold prey.

Image credit: Ra’ike
Rostrum cross-sections are circular to oval, where the latter may be oriented along the dorsal-ventral plane, or at right angles to this plane. The robust rostrum is the commonly preserved belemnite component, the more delicate phragmocone less so. It seems likely that the rostrum was part of the buoyancy apparatus in living forms.

Bottom: Waisted rostrum of the Upper Lias Acrocoelites. Photos courtesy of Annette Lokier, University of Derby.
Bottom right: The sketch of an idealised cross section of the rostrum, depicts progressive radial crystal growth (probably syntaxial) corresponding to the addition of successive calcite layers.
Other posts in this series
Bivalve shell morphology for sedimentologists
Gastropod shell morphology for sedimentologists
Brachiopod morphology for sedimentologists
Trilobite morphology for sedimentologists
Echinoderm morphology for sedimentologists
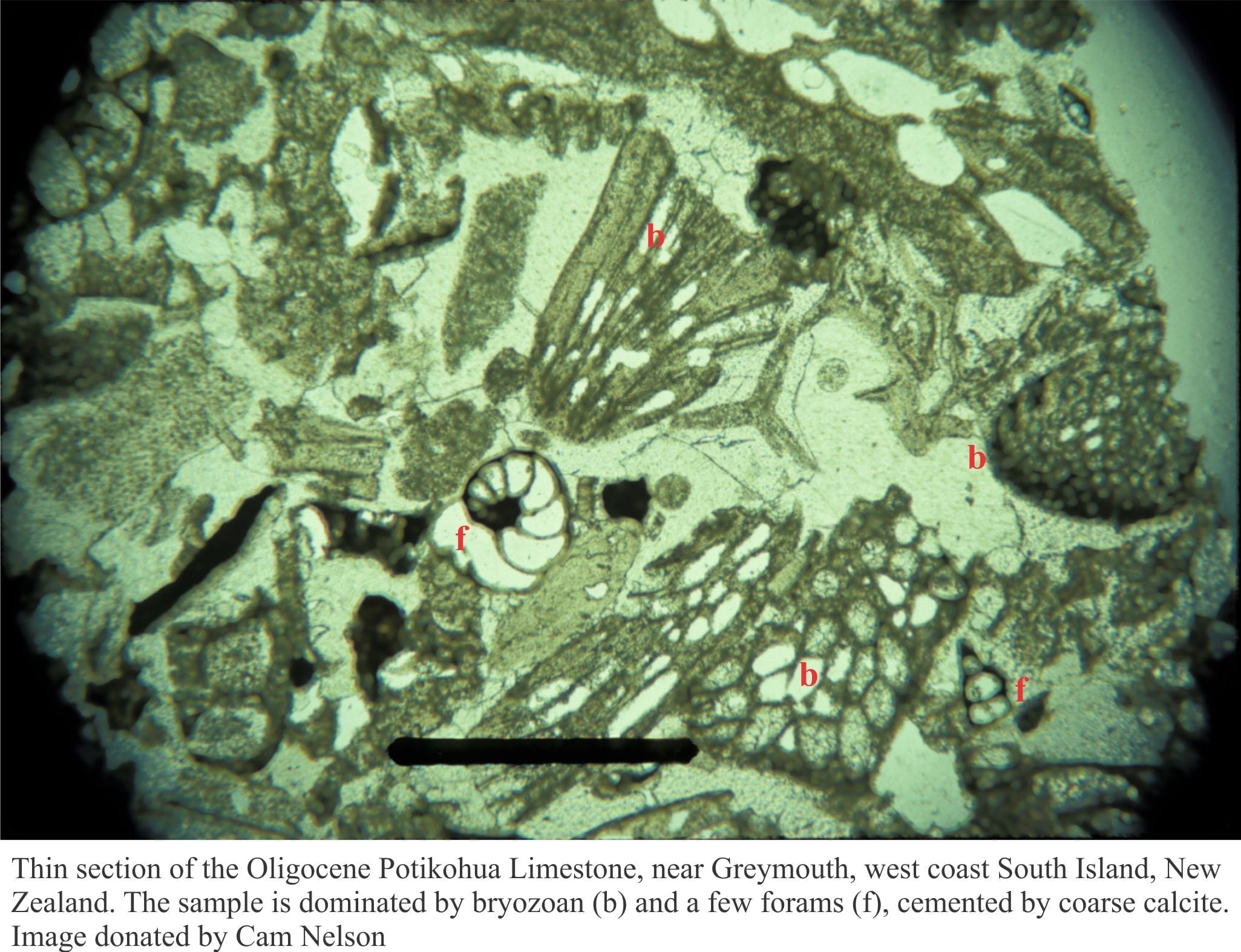
Carbonates in thin section: Molluscan bioclasts
Mineralogy of carbonates; skeletal grains
Mineralogy of carbonates; non-skeletal grains
Mineralogy of carbonates; lime mud
Mineralogy of carbonates; classification
Mineralogy of carbonates; carbonate factories
Mineralogy of carbonates; basic geochemistry
Mineralogy of carbonates; cements
Mineralogy of carbonates; sea floor diagenesis
Mineralogy of carbonates; Beachrock
Mineralogy of carbonates; deep sea diagenesis
Mineralogy of carbonates; meteoric hydrogeology
Mineralogy of carbonates; Karst
Mineralogy of carbonates; Burial diagenesis
Mineralogy of carbonates; Neomorphism